Review Article - (2015) Volume 4, Issue 4
Background: Since viral infections activate type I interferon (IFN) pathways and cause subsequent release of IFN-dependent proinflammatory chemokines/cytokines, the innate immune system plays an important role in the pathogenesis of lupus nephritis (LN) as well as systemic lupus erythematosus. With respect to this issue, mesangial chemokine/cytokine expressions via innate immunity reportedly play a pivotal role in the pathogenesis of LN. Also, it has been reported that pediatric-onset active LN is found as having more severe acute inflammation than that of adult cases. Therefore, viral or “pseudoviral” infections may be attributable, at least in part, to the pathogenesis of LN, especially in pediatric-onset cases.
Methods: So far, we have examined the toll-like receptor (TLR) 3 signaling cascades treated with polyinosinicpolycytidylic acid (poly IC), a synthetic analogue of viral dsRNA, that makes “pseudoviral” infection in cultured human mesangial cells (MCs).
Results: We found that the activation of mesangial TLR3 upregulated the expression of functional molecules acting as monocyte/macrophage and lymphocyte chemoattractants in MCs. Further, intense glomerular expressions of these functional molecules were observed in biopsy specimens from children with active LN.
Conclusion: These observations further support the implication of viral and “pseudoviral” immunity in the pathogenesis of active LN, especially in pediatric-onset cases.
Keywords: Innate immunity; Mesangial cells; Proinflammatory chemokines; Active lupus nephritis; Pseudoviral immunity; Toll-like receptor 3
Although lupus nephritis (LN) is one of the major clinical manifestations of systemic lupus erythematosus (SLE), the pathogenesis of active LN remains to be elucidated. Given the implication of innate immunity mediated by the activation of signaling cascades via Toll-like receptors (TLRs) in the pathogenesis of LN as well as SLE [1-4], the involvement of renal TLRs and their downstream signaling pathways have been studied so far [1,4-8]. Interestingly, activation of immune responses via TLRs can be induced by both infectious pathogens and non-infectious stimuli such as endogenous ligands. Since this mechanism, so-called “pseudoviral” immunity, may be possibly involved in the pathogenesis of LN [1-4], we have examined the TLR3 signaling cascades induced by polyinosinic-polycytidylic acid (poly IC), a synthetic analogue of viral dsRNA, which elicits a “pseudoviral” infection in normal human mesangial cells (MCs) in culture, and found that mesangial TLR3 activation increased the expression of downstream functional molecules acting as chemoattractants for inflammatory cells in MCs [5-12]. Since pediatric-onset active LN is reportedly found as having more severe acute inflammation than that of adult cases [13], we believe that viral or “pseudoviral” infections is attributable, at least in part, in the pathogenesis of the disease [1,4,6].
We discovered that IFN-β, synthesized de novo following TLR3 activation, may be a key molecule in the regional inflammatory cascades in MCs in our experimental setting [10-12]. With respect to this issue, we believe that in our experimental setting that utilized human MCs, de novo synthesized IFN-β acts, even partially, as an “autocrine” mediator in residual renal cells [5]. In contrast, IFN-α is probably released by infiltrating proinflammatory cells and acts as a “paracrine” mediator in various clinical and experimental settings, although this theory remains speculative. Recently, an interesting case of a so-called IFN-β nephropathy has been reported, which developed after a long-term IFN-β treatment for relapsing multiple sclerosis [14]. Thus, the regional role of IFN-β in MCs, especially in the case of a sustained exposure to this substance, remains to be properly elucidated. Furthermore, we confirmed intense glomerular expressions of IFN-β-dependent molecules in biopsy specimens from children with active LN (class III and IV in accordance with International Society of Nephrology/Renal Pathology Society 2003 classification for LN), whereas negative staining occurred in specimens from patients with other form of immunocomplex-mediated glomerulonephritis, such as IgA nephropathy (IgAN) or purpura nephritis (PN) [5,6,8,11,12]. These observations further support the implication of regional activation of TLR3/IFN-β signaling cascades in the pathogenesis of pediatric-onset active LN.
It has also been reported that glomerular expressions of TLRs (TLR 3,4,7 and 9) may play pivotal roles in the pathogenesis of LN and IgAN [1-3,15,16]. Since mammalian endogenous nucleic acids as well as bacterial DNA or viral RNA were reportedly recognized by TLR 7 and TLR 9, the generation of some autoantibodies may attribute to the activations of these receptors in selected patients with SLE and LN [3,16,17]. Also, SLE is well known as the prototype of systemic autoimmune disorders [3]. However, here, based on our recent findings, we focus on some aspects of the activation of TLR3 signaling pathways in active LN, which is possibly involved in the pathogenesis of the disease [1,4,6].
It has been reported the implications of several type I IFN-dependent transcripts in innate immunity, including human myxovirus resistance protein 1 (Mx1), and Mx1 reportedly acts as an inhibitor of RNA viral replication [18]. Although its implication in the pathogenesis of human SLE remains to be elucidated, it has been reported that measurement of Mx1 gene expression in peripheral blood cells from patients with LN may be a possible biomarker for therapeutic intervention of such patients [19]. However, the implication of mesangial expression of Mx1 in the pathogenesis of immunocomplex-mediated glomerulonephritis is still unclear [20].
Recently, we found that Mx1 expression was induced time- and dose-dependently by poly IC in MCs, and RNA interference (siRNA) against IFN-β effectively inhibited the expression [5]. In addition, intense mesangial Mx1 expression was confirmed in renal biopsy specimens obtained from pediatric patients with active LN (Class III and IV), whereas negative results were observed in specimens of patients with other form of immunocomplex-mediated active glomerulonephritis, such as IgAN or PN. Thus, we speculated that the activation of TLR3/IFN-β signaling pathways is responsible for mesangial expression of this IFN-dependent transcript [5,11]. However, we previously found that the cell viability and the downstream expressions of various IFN-stimulated genes were not influenced by Mx1 knockdown [21]. Thus, the detailed function of Mx1 remains to be determined in the future studies. Although mesangial expression of Mx1 may be a sequel of regional type I IFN activation, we believe that our novel finding may further support the theory of marked activation of viral and/or “pseudoviral” innate immunity in the pathogenesis of active LN [5].
CXCL1/GROα is a strong neutrophil chemoattractant cytokine and reportedly plays an important role in regional inflammatory reactions. It has been reported that stimulations by pathogenic anti-DNA antibodies and nucleosome-containing immune complexes induced mesangial expressions of CXCL1 and CXCL2 in murine models of LN [22].
With respect to this issue, we found that poly IC treatment induces time- and dose-dependently expression of CXCL1 in MCs. On the other hand, treatment of cells with poly IC/cationic lipid complex did not induce glomerular CXCL1 expression, although it apparently induced CXCL10 (also known as IP-10) expression [8]. Further, we found that siRNA-mediated knockdown of TLR3, but not cytosolic RNA recognition sensors, retinoic and-inducible gene-I (RIG-I) and melanoma differentiation-associated gene 5 (MDA5), suppressed poly IC-induced CXCL1 expression, indicating that TLR3 acts as a dominant receptor for this reaction. TLR3, RIG-I and MDA5 are known as pattern-recognition sensors for viral dsRNA [23]. Thus, mesangial expression of CXCL1 induced by poly IC may mainly depend on the endosomal TLR3 [8,12]. Also, we found mesangial CXCL1 immunoreactivity in the specimens from pediatric patients with active LN (Class III and IV) but a trace CXCL1 expression in IgAN specimens (Figure 1). Thus, our observation suggested that apparent activation of TLR3 signaling pathways exists in the glomerular lesions of patients with LN. Since CXCL1 has been reported to play as podocytotoxic [24] as well as a chemokine chemotactic to neutrophils, the implication of this cytokine is growing in the development of LN. On the other hand, it has been reported that mesangial CXCL2 expression induced by nucleosomes possibly plays a pivotal role in the development of LN in mice [25]. However, the detailed roles of expressions of mesangial CXCL1 and CXCL2 in the development of human LN remain to be elucidated. Further studies are needed.
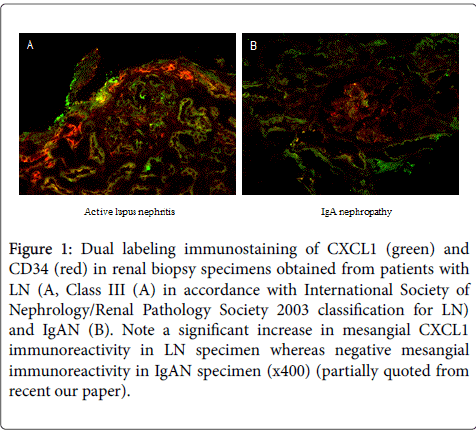
Figure 1: Dual labeling immunostaining of CXCL1 (green) and CD34 (red) in renal biopsy specimens obtained from patients with LN (A, Class III (A) in accordance with International Society of Nephrology/Renal Pathology Society 2003 classification for LN) and IgAN (B). Note a significant increase in mesangial CXCL1 immunoreactivity in LN specimen whereas negative mesangial immunoreactivity in IgAN specimen (x400) (partially quoted from recent our paper).
Although cross-talk of various proinflammatory chemokines/cytokines induced by TLRs activations play protective or deleterious roles in glomerular inflammation [8], we believe that TLR3 activation namely “pseudoviral” immunity in MCs [1] also plays an important role in the development of LN.
It has been reported that active SLE patients have both systemic and regional activation of proinflammatory cytokines [26,27]. Recently, Koenig et al. reported a marked increase in serum soluble receptor of tumor necrosis factor (TNF)-α in lupus patients with active stage, suggesting the presence of possible activation of TNF-α [26]. However, in the lupus models, TNF-α reportedly plays both beneficial and deleterious roles in the development of LN [27]. Therefore, the implication of regional TNF-α activation in the development of human LN remains to be elucidated.
We recently found that pretreatment of MCs with TNF-α of 1 ng/ml (possibly low dose) markedly enhanced the expression of CCL5 induced by poly IC [7]. We also confirmed pretreatment of MCs with TNF-α of 1 ng/ml enhanced the expression of IFN-β induced by poly IC, whereas TNF-α at the same dose alone did not induce the expression of IFN-β. Further, siRNA against IFN-β significantly reduced the expression of CCL5 induced by TNF-α followed by poly IC [7]. These results indicated that the synergistic induction of CCL5 in MCs can be achieved by the combined pretreatment of MCs with TNF-α (even at low dose?) followed by poly IC. Therefore, this experimental observation suggests that pre-existing regional activation of TNF-α results in the enhanced glomerular expression of CCL5 in patients with active SLE, which may be involved in the development of LN. In this experimental setting using MCs, an increased IFN-β due to pre-existing TNF-α activation may be a key mediator of enhanced expression of CCL5 [7].
In a clinical setting also, beneficial effect of TNF-α blockade of early induction treatment has previously been reported in selected patients with difficult LN [28]. Taking together, in active SLE patients, both pre-existing TNF-α activation and subsequent activation of the TLR3/IFN-β signaling may be involved in the development of LN [7].
Given the implication of “pseudoviral” immunity in the pathogenesis of LN [1], we believe that involvement of the TLR3/IFN-β signaling activations described here in MCs possibly contribute to mesangial inflammation (Figure 2), that may be involved in the pathogenesis of active LN. Since the treatment using corticosteroid depend on the inhibitory effect of IFN-β production and not on the activation of downstream cascades following de novo synthesized IFN-β [9,10], effective treatment strategies for the intervening in the TLR3/IFN-β cascades are desirable for the future therapeutic strategies for active LN.
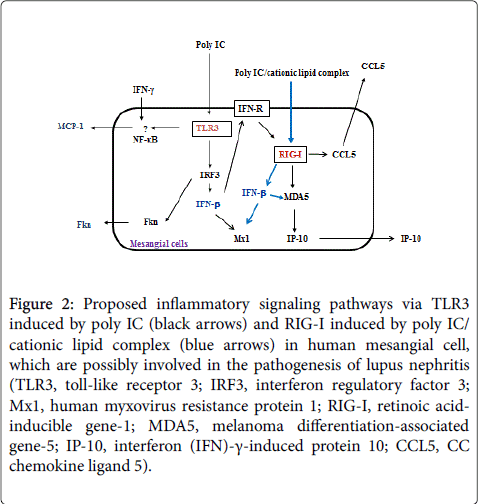
Figure 2: Proposed inflammatory signaling pathways via TLR3 induced by poly IC (black arrows) and RIG-I induced by poly IC/ cationic lipid complex (blue arrows) in human mesangial cell, which are possibly involved in the pathogenesis of lupus nephritis (TLR3, toll-like receptor 3; IRF3, interferon regulatory factor 3; Mx1, human myxovirus resistance protein 1; RIG-I, retinoic acidinducible gene-1; MDA5, melanoma differentiation-associated gene-5; IP-10, interferon (IFN)-γ-induced protein 10; CCL5, CC chemokine ligand 5).
Part of our research involving human subjects was approved by the ethics committee of Hirosaki University Graduate School of Medicine.
These studies were supported by Grants-in-Aid for Japan Society for Promotion of Science (JSPS KAKENHI Grant Number 25461615)."We thank Prof. Kensuke Joh, Department of Pathology, Tohoku University Graduate School of Medicine for performing dual immunostaining of biopsy specimens.